|
|
 |
|
Плауновидные карбона, что они кроме лепидодендрона и сигиллярии
Автор: donbassfossil
Рубрика: Обсуждение литературы
Это скорее реферат, нежели самостоятельно подготовленная публикация, однако, с моей точки зрения, эта информация довольно полезна тем, кто пытается разобраться в сути классификации и морфологии ископаемых плауновидных. Она основана на открытых публикациях на английском языке и также полезна тем, кто пока не освоил язык до нужного уровня.
Несмотря на огромное количество накопленной информации об ископаемых плауновидных, в нашем понимании их эволюции все еще остаются белые пятна. В Ероамерике плауновидные были представлены в карбоне, по крайней мере, четырьмя главными порядками, которые включают крупные древовидные формы лепидодендровых (Lepidodendrales) – лепидодендрон (Lepidodendron), диафородендрон (Diaphorodendron), лепидофлойоз ( Lepidophloios)и сигиллярия (Sigillaria), имеющих стигмариевидную корневую систему; более мелкие древовидные формы полушниковых (Isoetales) – в частности, Chaloneria, которая имеет клубневидную корневую систему и травянистые таксоны, с лигулой или без нее, включая селагинелловые (Selaginellales) и плауновые (Lycopodiales). Однако известно, в карбоне существовали и другие плауновидные, как, например, Oxroadia , Paurodendron (Selaginella), Paralycopodites(Anabathra) или недавно выделенный род Hizemodendron, которые были травянистыми или псевдотравянистыми формами, однако, продуцировали моноспорангиальную шишку наподобие лепидодендровых. О лепидодендронах, сигилляриях, лепидофлойозах и диафородендронах коротко было сказано в предыдущих публикациях, они же скромно представлены в моих фотогалереях. Здесь хотелось бы сказать пару слов о других членах этого «забавного семейства».

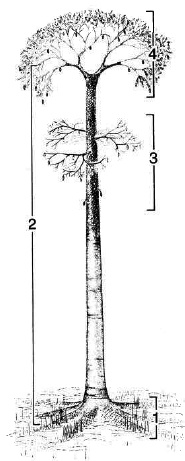
Древовидные лепидодендровые имеют общие характеристики архитектуры растения (“bauplan”), которые включают: 1 - радиально-симметричные ризоморфы (стигмарии), 2 - ствол (от корня до первой изотомии меристемы аписа), 3 - боковые ветви (или фертильные плодоножки) и 4 - ветви кроны (см. рис.).
Oxroadia
Травянистое плауновидное с дихотомически разветвляющимся стволом без выступающих над поверхностью листовых подушек (ЛП). Отсутствует вторичное утолщение, листья расположены спирально, имеют единственную центрально расположенную жилку и лигулу, листья не опадают, парихны отсутствуют.
Кстати, о лигуле. Гетероспоровые плауновидные имеют специфическое углубление на поверхности листа поблизости от места прикрепления, называемое лигулой. На отпечатках его можно видеть чуть выше листового рубца в пределах базы листа. Эти углубления развиваются в период, когда лист формируется, их роль – выделять особый секрет для увлажнения растущего листа.
Paralycopodites (Anabathra)
Имеет листовые подушки, которые по форме занимают промежуточное положение между лепидодендроном и лепидофлойозом. На больших диаметрах ствола ЛП ромбической или треугольной формы, скорее напоминающие лепидофлойоз, на которых располагались листовые рубцы (приблизительно по центру), имеющие три рубчика (проводящий пучок по центру и парихны по краям). Листья оставались все время прикрепленными к боковым ветвям, межлистовые расстояния были незначительными, лигула расположена на стволе очень близко над ЛП. Киль без перпендикулярных бороздок присутствует на нижней части ЛП. На малых диаметрах ветвей ЛП могут становиться вертикально вытянутыми, более походить на лепидодендрон.
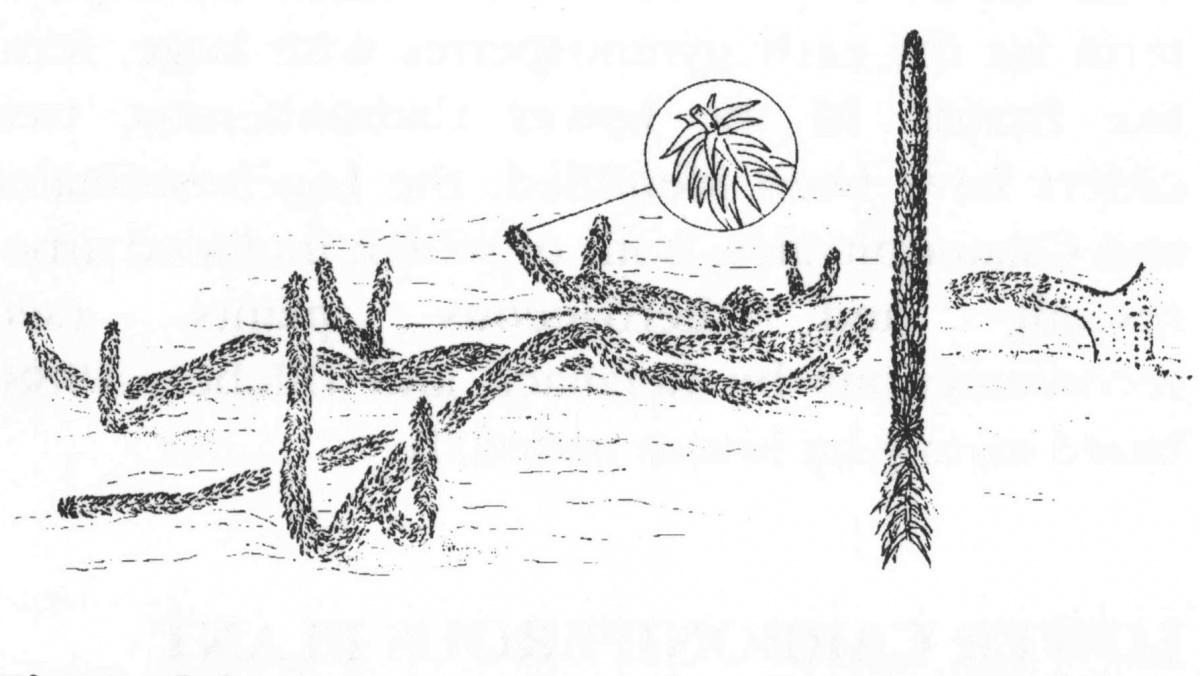
Хиземодендрон (Hizemodendron serratum) является представителем лепидодендровых, для которых характерно наличие лигулы, вторичной ксилемы, развитой перидермы, трехзоновой коры и стигмариевидной корневой системы. Предполагается, что этот тип морфологии растения развился путем уменьшения длины ствола изначально древовидного плаунообразного.
Архитектура Hizemodendron сходна с Oxroadia. Очень короткий (предположительно древесинный) ствол, возвышающийся над последовательно дихотомически ветвящимися стигмариями. Крона сформирована часто дихотомически ветвящимися под широким углом ветвями (как структурными, так и несущими спорофиллы), которые не являются древесинными и покрывают большую часть поверхности земли вокруг растения. Отличиями Hizemodendron от Oxroadia являются существенно больший размер растения и строгая анизотомия ветвления (что сближает его с Paurodendron (=Selaginella)). Таким образом, это было лежачее, карабкающееся растение, которое все же нельзя назвать распластавшимся. Ветви, расположенные вблизи ствола, имели извилистую форму, касаясь грунта изредка только некоторыми частями, что позволяло листьям активно фотосинтезировать.
Характерными особенностями формы листовых подушек (ЛП) являлись:
-Соотношение высота/ширина ЛП для малых диаметров ветвей менее 1
-Отсутствует механизм сохранения листьев
-Отсутствует верхний киль
-Верхняя часть ЛП без морщинок
-Отсутствует боковая линия, разделяющая верхнее поле ЛП от нижнего.
-Отсутствуют межлистовые парихны.
Взглянем на кладограмму плауновидных из работы W. DiMichele.
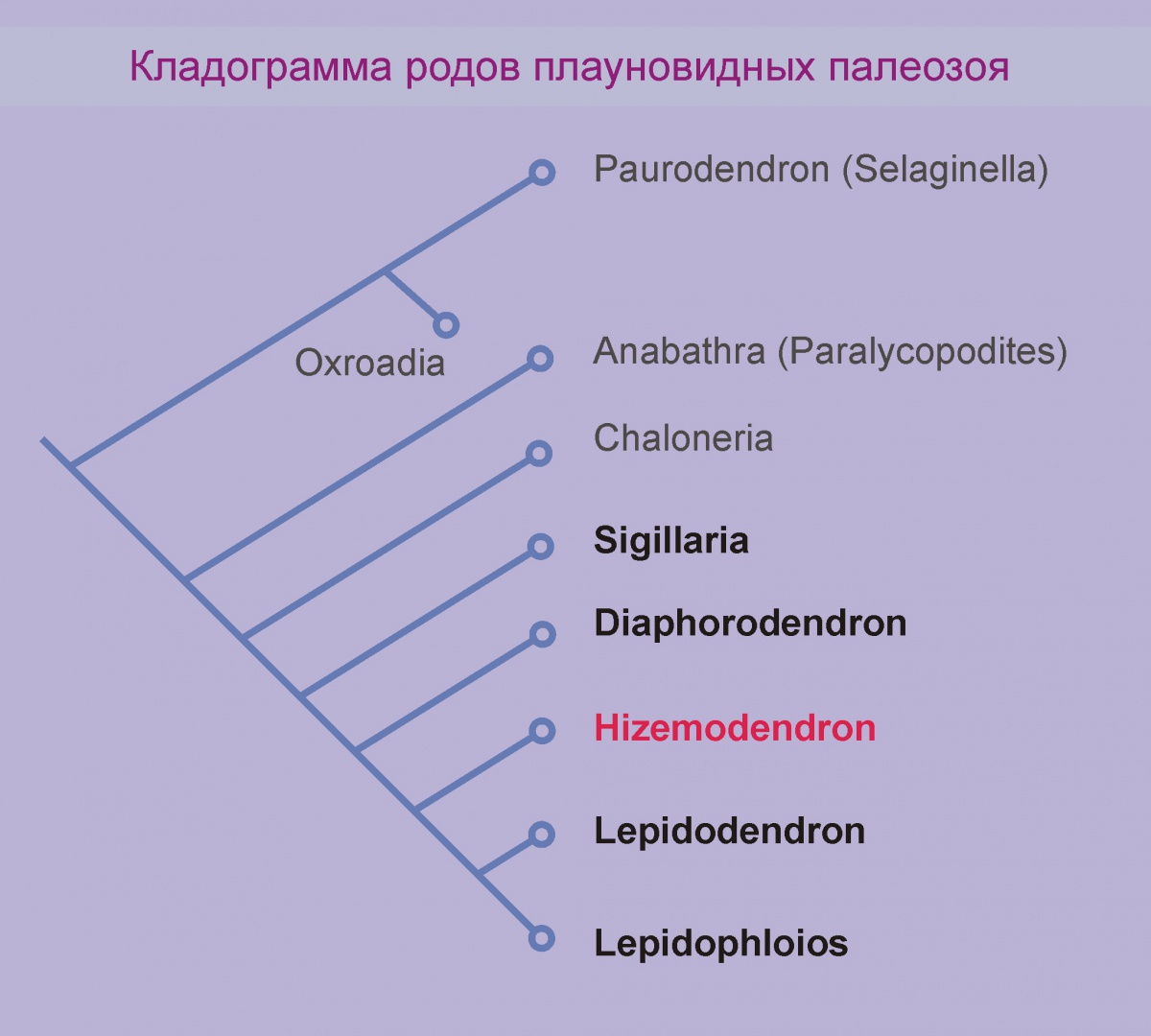
Кладограммами и соответствующим анализом занимается наука филогенетика.
Еще ссылки для интересующихся филогенетикой.
Емельянов А.Ф., Расницын А.П. Систематика, филогения, кладистика // Природа. – 1991. – № 7. – С. 26–37.
Становление современной филогенетики.
Те, кто слегка углублялся в палеоботанику и классическую систематику карбоновых плауновидных, спросит, а где же широко известная группа растений асоланус - пинакодендрон - циклостигма с формой отпечатков коры без листовых подушек? А не было таких растений, по мнению американских палеоботаников. Многочисленные отпечатки коры с листовыми рубцами в плоскости ствола и характерным волнообразным или диагональным орнаментом, находимые в разных частях света, являются просто различными формами декортикации коры другого плауновидного – Chaloneria cormosa с клубневидной корневой системой (а не стигмариями).
Chaloneria cormosa
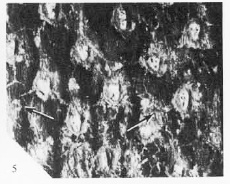
Исторически сложилось так, что многие палеоботаники полагали, что имеющийся палеонтологический материал, хоть по общему признанию и не полный, позволяет утверждать, что полушниковые (Isoetes ) являются венцом серии трансформационных преобразований в карбоне. Предполагалось, что, начиная с сигиллярии, у которой ветвей минимум, необходимость в разветвленной мощной корневой системе стигмариевого типа стала ослабевать и было положено начало переходу к клубневидной корневой системе и уменьшению общего размера растения. Плевромея (Pleuromeia) из триаса является промежуточным звеном в этой цепи преобразований, а конечное звено – Isoetes. Однако, открытие плауновидных карбона с клубневидной корневой системой, такого как Chaloneria, свидетельствует о том, что они не были результатом постепенного уменьшения размеров древовидных образцов, а существовали одновременно с ними.
Остаются не упомянутыми еще такие виды плаунообразных, как улодендрон (Ulodendron) и ботродендрон (Bothrodendron).
Что касается улодендрона, несмотря на то, что Томас (Thomas) выделял его в отдельный род, Jonker считал, что поскольку веточные рубцы, диаметрально противоположно расположенные на стволе - характерная черта лепидодендронов, лепидофлойозов и ботродендронов, то их, наверное, можно смело объединять с этими видовыми именами. DiMichele считает улодендрон близким родственником Paralycopodites(Anabathra).
Ботродендрон известен главным образом по отпечаткам коры. Если я не ошибаюсь, у Тейлора (первая из цитируемых работ в этой публикации) описана споровая шишка Bothrodendrostrobus, имеющая анатомическую сохранность. Этот образец интересен тем, что на нем видны два зародышевых придатка, выходящих за пределы споровой камеры. Один является первым побегом, второй – первым корнем. Таким образом стало понятно, что эмбриология лепидодендрона (Lepidocarpon) и ботродендрона (Bothrodendrostrobus) слишком разнятся и представляют собой две эволюционные ветви внутри плауновидных. Это было интерпретировано еще также и как то, что Bothrodendrostrobus (биполярное развитие) является производным от Lepidocarpon (монополярное).
Тем не менее, в кладограмме DiMichele ботродендрон не показан, а в публикации о Chaloneria cormosa (последняя из цитируемых работ в этой публикации) утверждается, что среди образцов этого растения были найдены и очень напоминающие отпечатки коры ботродендрона. Так что вопрос пока остается открытым.
Источники текстового описания и иллюстраций:
THOMAS N. TAYLOR at al., PALEOBOTANY, The Biology and Evolution of Fossil Plants, Second Edition. 2009.
Department of Ecology and Evolutionary Biology and Natural History Museum and Biodiversity Research Center, The University of Kansas, Lawrence, Kansas.
Bateman , R. M. , and W. A. DiMichele. 1991. Hizemodendron , gen. nov., a pseudoherbaceous segregate of Lepidodendron (Pennsylvanian): Phylogenetic context for evolutionary changes in lycopsid growth architecture . American Society of Plant Taxonomists 16: 195 – 205.
K.L. Alvin, A New Fertile Lycopod from the Lower Carboniferous of Scotland, Palaeontology, Vol.8, Part 2, 1965, pp. 281-293.
William A. DiMichele. Paralycopodites Morey & Morey, from the Carboniferous of Euramerica. A Reassessment of Generic Affinities and Evolution of "Lepidodendron" brevifolium Williamson. American Journal of Botany, Vol. 67, No. 10 (Nov. - Dec, 1980), 1466-1476.
K. B. Pigg, G. W. Rothwell. Cortical development in Chaloneria cormosa (Isoetales), and the biological derivation of compressed lycophyte decortication taxa. Palaeontology, Vol. 28, Part 3, 1985, pp. 545-553.
|
|
Ключевые слова (тэги): плауновидные, филогенетика
 Добавить в избранное
Добавить в избранное
|
Постоянный адрес публикации: |
HTML-ссылка на публикацию: |
Публикация создана 12 января 2012 года
Если вы хотите оставить комментарий,
вам нужно зарегистрироваться (или авторизоваться, если вы уже зарегистрированы)
Комментарии:
|
<a href="http://ammonit.ru/text/338.htm#53432">http://ammonit.ru/text/338.htm#53432</a> |
|
|||||
Интересное дополнение! | ||||||
|
комментарий 1 уровня |
Комментарий создан 16 января 2012 года в 13:48:06 |
|||||
 Добавить в избранное
Добавить в избранное 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
|
<a href="http://ammonit.ru/text/338.htm#53519">http://ammonit.ru/text/338.htm#53519</a> |
|
|||||
Создается впечатление, что чем древнее флора и фауна, тем строже у них геометрические формы... | ||||||
|
комментарий 1 уровня |
Комментарий создан 16 января 2012 года в 23:47:56 |
|||||