|
|
 |
|
Антидихотомия
Автор: donbassfossil
Рубрика: Палеонтологические публикации
Так уж сложилось, что отпечатки плауновидных карбона – одни из самых часто встречаемых в местах моих поисков. В одном месте было найдено целое «месторождение» ботродендронов. Ботродендрон – типичный, но редкий представитель плауновидных, отличительной особенностью которого являются рубцы боковых ветвей на стволе, выстроенных взаимно противоположными вертикальными рядами.
Принято считать, что все плауновидные ветвятся дихотомически. То есть, активный участок роста верхушки ствола или побега (верхушечная апикальная меристема) все время делится надвое. Если деление равнозначное – изотомическая дихотомия, если нет – анизотомическая. Если сказать точнее, для плауновидных характерно дихоподиальное ветвление. "Анизотомия переходит в дихоподиальное ветвление в результате резко неравного развития двух дочерних ветвей, когда одна из них перевершинивает другую. Процесс такого перевершинивания приводит к возникновению дихоподия, внешне напоминающего так называемый симподий цветковых растений, с его зигзагообразной главной осью. Дихоподиальное ветвление было характерно для примитивного девонского рода астероксилон (Asteroxylon), оно имеет место у многих плаунов и селагинелл, у некоторых древнейших хвощевидных и у многих папоротниковидных" - цитата из академического словаря в интернет.
Все эти рассуждения справедливы для тонких облиственных ветвей лепидодендрона или ботродендрона.

Что же касается боковых ветвей, характерные рубцы которых рядами остаются на стволах ботродендронов, тут не все сходится по следующим соображениям.
Начнем с работы Bateman 1994, которая предлагает для всех древовидных плаунообразных обязательное наличие А - корня (различной формы), В – ствола (большого или малого), С – кроны (хорошо или слабо развитой) и D - боковых ветвей ствола и кроны, которые могут присутствовать, а могут - нет. Причем, для сигиллярии и лепидофлойоза боковые ветви присутствуют в редуцированной форме побегов, несущих споровые шишки.
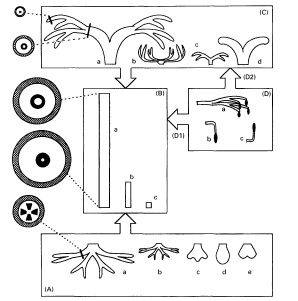
Вот, что он пишет далее:
"Латеральные ветви - наиболее проблематичные элементы морфологии ствола плауновидных и требуют дополнительного обсуждения. В отличие от других элементов структуры они могут занимать различное положение на стволе: либо до, либо после первой изотомии апикальной меристемы ствола [т.е. главной дихотомической вилки ствола, за которой формируется крона - прим. авт.]. Те боковые ветви, которые размещаются на ветвях кроны, являются более мелкими по сравнению с боковыми ветвями, присоединенными непосредственно к стволу. Боковые ветви разделены на два субмодуля, которые гомологичны структурно, а не по месту размещения. Оба субмодуля непременно производят спороносные шишки, хотя, к примеру, боковая ветвь сигиллярии состоит только из шишки плюс плодоножка. Гомологичность неразветвленных плодоножек и многократно дихотомирующих боковых ветвей подтверждается тем фактом, что нет ни одного из известных ризоморфных ликопсид, которые бы имели как боковые ветви, так и плодоножки прямо размещенные на стволе".
Примеры рубцов от редуцированных боковых ветвей для лепидофлойоза и сигиллярии показаны ниже (первое фото находки лепидофлойоза из моей коллекции, вторая – сигиллярия из известной коллекции Кидстона). Для сигиллярии характерно появление веточных рубцов (напоминаю, что это тонкие редуцированные ветви, по сути плодоножки для шишек) между вертикальными рядами листовых подушек. Отчего я по аналогии и делаю ниже вывод о "прорастании" боковых ветвей сквозь уже сформировавшуюся кору ствола ботродендрона.


Боковые (или, по-другому, латеральные стволовые) ветви всегда нарушали привычные представления об исключительно дихотомическом ветвлении лепидодендронов, особенно после того, как были найдены странные веточные рубцы на стволах, например, Lepidodendron veltheimi (фото из коллекции Кидстон).

или Lepidodendron nathorsti (взято оттуда же)

В прошлом году появилась публикация ДиМайкла (DiMichele W. at al. 2013) о найденном диафородендроне (точнее, его разновидности Synchysidendron) с боковыми стволовыми ветвями, подобно тем, которые характерны и для ботродендрона. В этой работе автор говорит о строгой билатеральной симметрии ствола Synchysidendron. Таким образом, исходя из такой системы разделения апекса, мы имеем разделение на три участка верхушки ствола (ствол и два симметрично расположенных побега боковых ветвей), но тогда, какая же это дихотомия? Возникает вопрос.
С другой стороны, о какой дихотомии при формировании боковых ветвей может идти речь в случае с лепидофлойозом или сигиллярией, когда на стволе возникает целый пояс плотно расположенных ветвей?
В уже цитировавшейся ранее работе Opluštil S. 2010 мы снова видим, что на дихотомически ветвящихся кронах Lepidodendron mannebachense появляются несущие шишки побеги, однако они также не являются результатом изотомического разделения, автор говорит о псевдо-моноподиальном побеге. Т.е. побег не является результатом дихотомии, но и моноподиальным его назвать автор не решается.

Ветви Lepidodendron mannebachense с рубцами фертильных побегов из моей коллекции.
Автор Opluštil S. 2010 для несущих споровые шишки веток предлагает псевдо-моноподиальные ответвления от ветвей кроны лепидодендрона (которые сформированы классической дихотомией). На фото ниже вы можете видеть такую ветвь с остатками псевдо-моноподиальных ответвлений (показаны стрелками) из его работы.

А теперь я размещаю ниже схему с образца Линдси (Lindsey M. 1915) из ее же работы по ботродендрону.
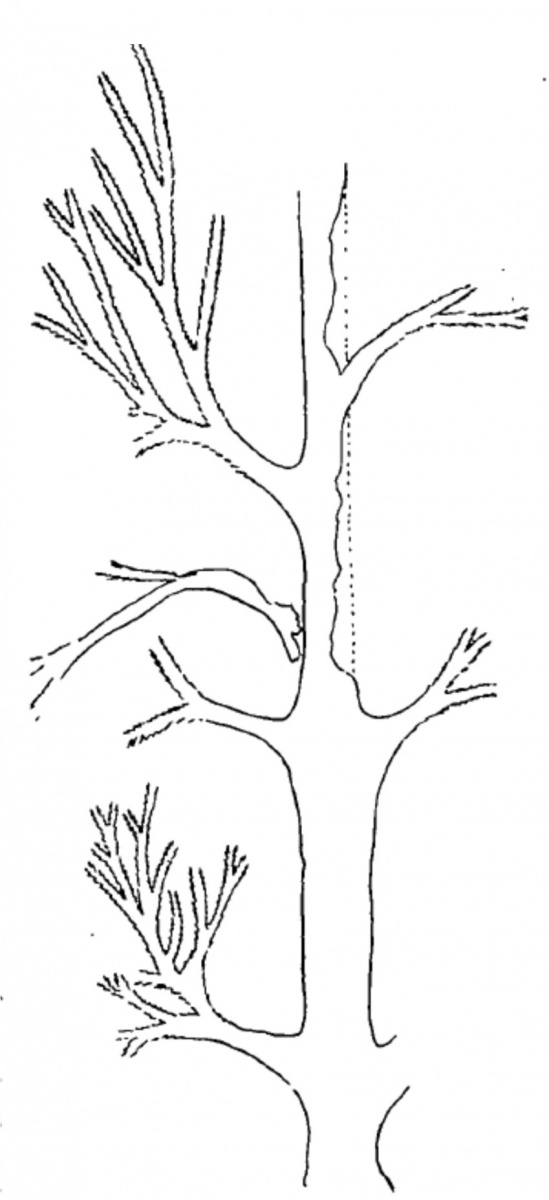
Ничего не напоминает? По-моему, один и тот же "почерк", очевидная гомология боковых ветвей у разных родов. В первом случае для боковых ветвей кроны, во втором - для латеральных стволовых ветвей. И те, и другие - могут нести стробилы.
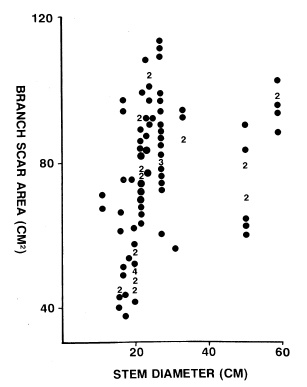
Теперь обратим взоры на уже представлявшуюся ранее в моих публикациях работу Wnuk C. 1989 по ботродендронам, где автор проанализировал многочисленные образцы стволов ботродендрона с веточными рубцами. Если боковые ветви появляются в результате дихотомического разделения вершины ствола (точнее, дихоподиального, с перевершиниванием), то должна прослеживаться четкая зависимость: чем толще ствол, тем больше размер веточного рубца. Просто потому, что ствол, разделившись на два побега, продолжает расти и увеличиваться в диаметре, но и образовавшаяся ветвь не замирает, а после ее отбрасывания, должен увеличиваться в размерах сам рубец. Обратите внимание на ниже расположенную диаграмму из работы Криса Внака 1989. На ней не прослеживается никакая четкая зависимость между диаметром ствола и площадью веточного рубца. Для одного и того же диаметра показаны площади рубцов в широком диапазоне разнообразия. Снова вопрос.
Предполагаю, что боковые стволовые ветви у всех плауновидных, в том числе ботродендрона, сигиллярии, лепидофлойоза и лепидодендрона маннебаченсе (псевдо-моноподиальные) появляются не в результате анизотомиии, а благодаря иному механизму (не связан с апикальной меристемой). Возможно, благодаря интеркалярным меристемам, осуществляющим вставочный рост. Они возникают уже после формирования ствола наподобие побегов от спящих почек современных деревьев. Постановка вопроса, конечно, носит весьма дискуссионный характер и может вызвать неприятие у палеоботаников. У кого есть другие соображения, с удовольствием выслушаю.
Справочно по видам меристем, ответственных за рост и ветвление растения, и типам ветвления:
В соответствии с расположением, помимо апикальных меристем, локализованных на кончиках осевых органов — стебля и корня, выделяют меристемы:
- латеральные (боковые) меристемы расположены внутри осевых органов и осуществляют их утолщение. К латеральным меристемам относят камбий, добавочный камбий и феллоген.
- краевая (маргинальная) меристема даёт начало листовой пластинке.
- интеркалярные меристемы осуществляют вставочный рост. За счёт их деятельности удлиняются междоузлия на ранних этапах развития побега, развиваются черешки листьев. Принципиально они отличаются от прочих меристем тем, что, помимо недифференцированных клеток, в них существуют и клетки на разных стадиях дифференциации. Поэтому корректнее здесь говорить не об интеркалярных меристемах, а о зонах интеркалярного (вставочного) роста.
Типы ветвления
Поскольку побег является осевым органом, у него присутствует апикальная меристема, которая обеспечивает неограниченное нарастание в длину. Такой рост сопровождается более или менее регулярным ветвлением побега. У низших растений в результате ветвления возникает разветвлённый таллом (слоевище), у высших — образуются системы побегов и корней. Ветвление позволяет многократно увеличивать фотосинтетическую поверхность и обеспечивать растение органическими веществами.
У различных растений наблюдается несколько типов ветвления: дихотомическое, моноподиальное, симподиальное. При дихотомическом ветвлении конус нарастания расщепляется надвое. При моноподиальном ветвлении верхушечная почка функционирует в течение всей жизни, образуя главный побег (ось первого порядка), на котором в акропетальной последовательности развиваются оси второго порядка, на них — третьего порядка и т.д. При симподиальном ветвлении одна из верхних пазушных почек образует ось второго порядка, которая растёт в том же направлении, что и ось первого порядка, смещая в сторону её отмирающую часть.
Справочные материалы взяты из интернета.
В итоге, моя очередная версия габитуса ботродендрона

Использованная литература:
Bateman R. 1994. Evolutionary-developmental change in the growth architecture of fossil rhizomorphic lycopsids: scenarios constructed on cladisticfoundations. Biol.Rev. Vol. 69, pp. 527-597.
DiMichele W., Elrick S. and R. Bateman. 2013. Growth habit of the Late Paleozoic rhizomrphic tree-lycopsid family Diaphorodendraceae: phylogenetic, evolutionary and paleoecological significance, American Journal of Botany 100(8): P. 1604–1625.
Lindsey M. 1915. The branching and branch shedding of Bothrodendron. Annals of Botany. Vol. 29. P. 223–230.
Opluštil S. 2010. Contribution to knowledge on ontogenetic developmental stages of Lepidodendron mannebachense Presl, 1838,, Bulletin of Geosciences, volume 85, issue 2; P. 303 — 316.
Wnuk C. 1989. Ontogeny and paleoecology of the middle Pennsylvanian arborescent lycopod Bothrodendron punctatum. Amer. J. Bot. Vol. 76 (7). P. 966–980.
Ключевые слова (тэги): ботродендрон, дихотомия
 Добавить в избранное
Добавить в избранное
|
Постоянный адрес публикации: |
HTML-ссылка на публикацию: |
Публикация создана 13 июля 2014 года
Если вы хотите оставить комментарий,
вам нужно зарегистрироваться (или авторизоваться, если вы уже зарегистрированы)
Комментарии:
|
<a href="http://ammonit.ru/text/1312.htm#194317">http://ammonit.ru/text/1312.htm#194317</a> |
|
|||||
Интересная работа. Спасибо. | ||||||
|
комментарий 1 уровня |
Комментарий создан 13 июля 2014 года в 17:00:56 |
|||||
 Добавить в избранное
Добавить в избранное 


|
<a href="http://ammonit.ru/text/1312.htm#194320">http://ammonit.ru/text/1312.htm#194320</a> |
|
|||||
Спасибо за внимание. | ||||||
|
комментарий 2 уровня |
Комментарий создан 13 июля 2014 года в 17:16:44 |
|||||

|
<a href="http://ammonit.ru/text/1312.htm#194350">http://ammonit.ru/text/1312.htm#194350</a> |
|
|||||
Интересно и познавательно, полезная публикация. | ||||||
|
комментарий 1 уровня |
Комментарий создан 13 июля 2014 года в 21:04:36 |
|||||
|
<a href="http://ammonit.ru/text/1312.htm#194360">http://ammonit.ru/text/1312.htm#194360</a> |
|
|||||
Благодарю Вас, Андрей! | ||||||
|
комментарий 2 уровня |
Комментарий создан 13 июля 2014 года в 21:37:01 |
|||||
|
<a href="http://ammonit.ru/text/1312.htm#194446">http://ammonit.ru/text/1312.htm#194446</a> |
|
|||||
Большое спасибо за интересную публикацию!
| ||||||
|
комментарий 1 уровня |
Комментарий создан 14 июля 2014 года в 14:51:02 |
|||||

|
<a href="http://ammonit.ru/text/1312.htm#194535">http://ammonit.ru/text/1312.htm#194535</a> |
|
|||||
Спасибо, Сергей! Публикация периодически дополняется и расширяется. | ||||||
|
комментарий 2 уровня |
Комментарий создан 15 июля 2014 года в 16:34:33 |
|||||
|
<a href="http://ammonit.ru/text/1312.htm#194486">http://ammonit.ru/text/1312.htm#194486</a> |
|
|||||
Все по полочкам ,спасибо за публикацию. | ||||||
|
комментарий 1 уровня |
Комментарий создан 14 июля 2014 года в 22:20:59 |
|||||

|
<a href="http://ammonit.ru/text/1312.htm#194487">http://ammonit.ru/text/1312.htm#194487</a> |
|
|||||
Спасибо, Юрий! | ||||||
|
комментарий 2 уровня |
Комментарий создан 14 июля 2014 года в 22:25:54 |
|||||