|
|
 |
|
Диморфизм аммонитов. Часть 2.
Автор: Alex
Рубрика: Палеонтологические публикации
Уважаемые коллеги! Перед вами вторая часть публикации о диморфизме аммонитов. Первая часть расположена здесь: Диморфизм аммонитов. Часть 1. Мегаконхи и миниконхи – кто они такие?. В первой части я рассказал о том, что среди аммонитов существовали два морфологических типа - микроконхи и макроконхи. Однако, среди микроконхов и макроконхов каждого вида часто встречаются как мелкие, так и крупные особи, причем иногда между ними нет переходных по размеру форм. В этой части рассказывается о причинах возникновения такой ситуации. Надеюсь, что статья не получилась слишком занудной и что дискуссия в комментариях к ней окажется столь же интересной, как и обсуждение первой части публикации.
Объективно в древних морях существовало только два морфотипа раковин аммонитов – микроконхи и макроконхи. И их отличия заключались не только в размерах, это был целый комплекс признаков – форма взрослого устья, угол навивания раковины, скульптура взрослых оборотов, учащение перегородок перед жилой камерой и т.д.. Не у всех родов аммонитов присутствовали все эти признаки и не у всех они проявлялись одинаково. Например, в волжском веке практически у всех аммонитов были одинаковые устья – без всяких боковых ушек. В меловом периоде «классические» длинные ушки встречались очень редко, но выросты на устьях присутствовали у многих форм, причем не только в отряде аммонитид, но и у литоцератид и филлоцератид. Но в общем, на протяжении практически двух периодов мезозоя у всех аммонитов существовали два морфотипа, различающихся между собой по целому комплексу признаков.
Первый морфотип – микроконхи – это относительно небольшие раковины, с постоянным (как правило), не меняющимся практически до самого конца взрослой жилой камеры типом скульптуры (довольно часто бывает, что скульптура усиливается, но общий ее тип не меняется), с постоянным (чаще всего) углом навивания раковины и с разнообразными выростами на терминальном устье (которых может и не быть).

Фото Два Indosphinctes nikitinoensis, автор Alex
Второй тип – макроконхи – относительно более крупные раковины, скульптура которых на взрослой жилой камере заметно менялась, как правило сглаживалась или заметно упрощалась, с ровным краем взрослого устья, часто с изменением угла навивания (с раскручиванием раковины) на последней жилой камере.

Фото Макроконх Indosphinctes wischniakoffi с устьем, автор Alex
И это всё - только два морфотипа. Все остальные варианты были выделены субъективно и говорить о том, что среди аммонитов были не только микроконхи и макроконхи, но и миниконхи с мегаконхами так же корректно, как и заявить, что среди людей кроме мужчин и женщин есть малорослые мужички и крупные дамы. Да, они есть, но малорослость и крупные габариты вторичны по сравнению с принадлежностью к полу, кроме того, рост, считающийся низким в одной популяции, в другой может быть средним, а то и выше среднего.
Тем не менее, сами аммониты из обеих субъективно выделенных групп – и «миниконхи» и «мегаконхи» все-таки существовали реально и об этом стоит поговорить отдельно.
Выше я обещал рассказать о теории полиморфизма развития, предложенной Матыем. Предварить этот рассказ нужно цитатой из книги известнейшего специалиста по современным головоногим, К.Н. Несиса: «У командорского кальмара имеются две сезонные нерестовые группировки - весенне-летняя и осенне-зимняя, впрочем, более вероятно, что даже три — отдельно осенняя и зимняя. Кальмары, родившиеся летом, созревают при длине мантии приблизительно на 2 см меньше, чем родившиеся осенью. В пределах каждой группировки выделяются раннеспелые и позднеспелые кальмары, немного различающиеся по размерам.» [6].
Именно на современных головоногих моллюсков, точнее, на кальмаров, обратил свое внимание Матый. Дело в том, что изначально многие как сторонники, так и противники теории полового диморфизма считали, что конечный размер раковин макроконхов и микроконхов постоянен для каждого вида. Откуда возникло это заблуждение сказать трудно. Между тем, наблюдения свидетельствовали об обратном – внутри одного вида разница в размерах между самыми маленькими и самыми крупными микроконхами могла быть больше, чем между крупными микроконхами и мелкими макроконхами. И ученые стали искать причину этого явления. Ответ подсказали современные головоногие. У осьминогов и кальмаров (именно к ним, а не к наутилусам были наиболее близки аммониты) существуют так называемые сезонные группировки. То есть, моллюски, родившиеся в разные сезоны, растут с разной скоростью и достигают полового созревания в разное время и при различной длине тела. А с наступлением полового созревания рост головоногих обычно заканчивается.
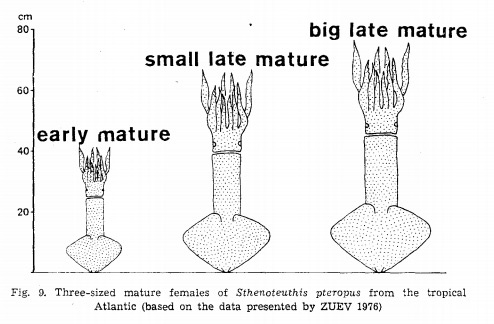
Разница в размерах взрослых самок кальмара Sthenoteuthis pteropus. Самые маленькие и самые крупные отличаются в два раза. Рисунок из публикации B.A. Matyia "Developmental polymorphism in Oxfordian ammonites"
К.Н. Несис приводит данные, что у некоторых кальмаров самки, родившиеся весной, уже следующим летом или осенью достигают половой зрелости, размножаются и умирают. То есть зимуют они лишь раз в жизни. А такие же самки, но родившиеся осенью, растут в течение всего следующего года, зимуют дважды и созревают только весной или летом на второй год жизни. По размерам и массе тела такие самки могут различаться в четыре раза, а то и больше.
Матый был первым палеонтологом, который экстраполировал эти данные на аммонитов. Таким образом, получила объяснение «тахигеронтность» и «брадигеронтность» аммонитов. Среди как микроконхов, так и макроконхов были быстровзрослеющие и медленновзрослеющие группы аммонитов – в зависимости от времени их появления на свет.
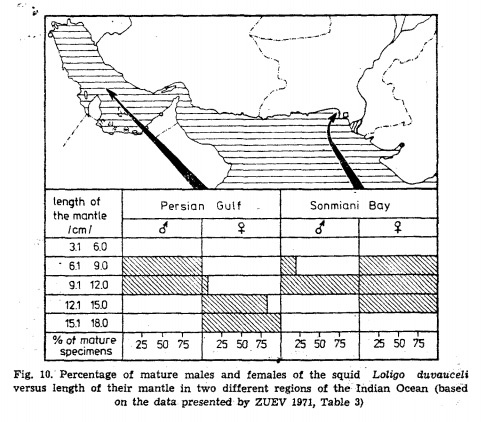
Разница в размерах самцов и самок в разных популяциях кальмара Loligo duvaucdi. Рисунок из публикации B.A. Matyia "Developmental polymorphism in Oxfordian ammonites"
Но Матый отрицал теорию полового диморфизма – он считал, что аммониты были удивительными животными, которые, в зависимости от момента достижения полового созревания, реализовывали различные морфологические типы. То есть если аммонит рос быстро, он отращивал ушки и жил с ними всю жизнь, а если рос медленно, то прекрасно обходился без всяких ушек, зато потом избавлялся от скульптуры на жилой камере. В дальнейшем эти аммониты как-то распределялись по местам обитания, а самцы и самки у них между собой если и различались, то крайне слабо.
Представить подобную картину конечно можно, но лишь на кратком участке истории живых организмов. В дальнейшем подобные виды неизбежно разделились бы на два независимых вида и разошлись бы навсегда на просторах эволюции. Но у аммонитов (причем в разных семействах) микроконхи и макроконхи существовали на протяжении десятков миллионов лет, совместно эволюционировали и не разделялись. Кроме того, Матый исходил из того, что любой макроконх «включает в себя» микроконх, то есть до какого-то момента они растут одинаково, просто микроконх останавливается, а макроконх продолжает расти. Такая ситуация действительно наблюдается у многих, но далеко не у всех аммонитов. У многих видов микро- и макроконхи различаются очень существенно уже на ранних оборотах и молодой микроконх на макроконха того же возраста может быть уже совсем не похож (как минимум по толщине, а иногда и по скульптуре). У каждого из них свой путь онтогенеза (индивидуального развития), но совместный эволюционный путь – их потомки также будут микроконхом и макроконхом одного вида, а не превратятся в разные биологические виды (хотя палеонтологи и могу записать их в разные формальные рода и виды).
То есть, теория полиморфизма развития в чистом виде не выдерживает критики. Представить себе аммонитов, реализующих практически произвольно один из двух морфотипов также сложно, как представить себе малька рыбы, который может вырасти либо в сома, либо в щуку в зависимости от темпов роста. Сам же Матый приводит в своих примерах современных головоногих лишь различия в размерах, а вовсе не в форме тела или образе жизни.

Фото Микроконхи Quenstedtoceras взрослели по-разному, автор Alex
Но внутри теории полового диморфизма представленная Матыем теория работает прекрасно, объясняя различия в размерах между крупными и мелкими самцами аммонитов (типичными микроконхами и «миниконхами») и крупными и мелкими самками («мегаконхами» и типичными макроконхами).
Интересно, что постулат Матыя, считавшего вслед за Тейссере микроконхов рано созревающими аммонитами, а макроконхов – поздно созревающими, тоже не противоречит истине. К аналогичным выводам пришел и другой противник теории полового диморфизма – А.Н. Иванов. Он считал, что микроконхи произошли от макроконхов путем неотении. В узком понимании этого термина неотения – приобретение способности к размножению на личиночных стадиях, а в широком – размножение на ранних этапах развития. Именно это и имел в виду Иванов. И он был, по-видимому, прав.
То есть тут конечно нужно уточнение – микроконхи произошли не от макроконхов, самцы не могли произойти от самок, нет, просто изначально самцы и самки могли расти примерно с одинаковой скоростью и достигать примерно одинакового размера. Такая ситуация наблюдается у современных наутилусов. Эти аммониты в стиле «унисекс» не обязательно во всем соответствовали морфотипу макроконхов, но они, скорее всего, были похожи на макроконхов по скорости роста и развития.
Нужно заметить, что у головоногих моллюсков, несмотря на то, что они существа довольно высокоразвитые, нет никакого взаимодействия между родителями и их детьми. Осьминоги и кальмары ничему не учат своих детенышей, роль самки ограничивается охраной кладки, а у многих видов и самцы и самки погибают сразу после нереста. То есть головоногим не нужно воспитывать свое потомство и им ничего не мешает максимально «выложиться по полной» в одном единственном в жизни акте размножения и после него умереть, оставив столько потомков, сколько сможет создать организм до своего полного истощения.
Конечно, для этого нужен быстрый обмен веществ и высокая скорость роста. Но колеоидеи, к которым относятся кальмары и осьминоги и к которым, по-видимому, были близки аммониты, растут очень быстро. Несис пишет, что «растут осьминоги быстро. Привес достигает 1%, а при обильном кормлении (в аквариуме) даже 1.8% веса тела в сутки. Летом на каждый килограмм съеденной пищи осьминог дает до 750 г привеса. Такое даже цыпленку-бройлеру не под силу! Зимой и весной рост замедляется. К концу первого года жизни масса осьминога достигает 100-130 г, второго - 1-2 кг, третьего - до 10 кг.».
Современный наутилус в этом ряду исключение, он строит свою раковину медленно, но каракатицы, к примеру, формируют свой внутренний скелет (кальцитовую пластинку сепион, по сути фрагмокон) очень быстро, за год-два. Вероятно, и аммониты формировали свою раковину за такой же срок.
Как это связано с быстро растущими самцами? Очень просто. Такие быстрорастущие головоногие включаются в своеобразную жизненную гонку – им нужно как можно быстрее, в прямом смысле слова жертвуя собственной жизнью, размножится и оставить как можно больше потомков. ««Жить быстро и умереть молодым!» — вот их стратегия.» - говорит о головоногих Несис.
Для наглядности проведем небольшой мысленный эксперимент - представим себе, что в каком-то условном виде аммонитов самцы и самки до момента полового созревания растут два года. Предположим, у нас есть 10 самцов этого вида, родившихся одновременно. Один из этих самцов оказывается мутантом, который был быстрее остальных и созрел за год. Пока все его сестры и 9 братьев созревают, он успевает к нересту аммонитов, родившихся на год раньше, и оставляет потомство.
У большинства головоногих нет сложного турнирного поведения самцов, которое бы мешало всякой мелочи получить доступ к самкам. А даже у тех видов, у которых такое поведение есть, у маленьких самцов всегда есть шанс пробраться к самке и сделать свое дело пока крупные самцы выясняют отношения. К.Н. Несис описывал такие ситуации у современных каракатиц. Так что наш мутантный аммонитик вполне мог оставить потомство, даже будучи вдвое меньше остальных самцов.
Для простоты картины предположим, что инкубация яиц происходит очень быстро и скоро в океане появляются детеныши этого самца, из которых половина наследует его скорость роста. Предположим, что из сотен «мальков» до взрослого состояния доживут немногие. Опять же для простоты картины пусть их будет 30 – по 15 самцов и самок, из них 7 самцов успеют созреть к следующему году. Кстати, чем быстрее они созревают, тем меньше вероятность, что многочисленные хищники просто не дадут им дожить до момента размножения.
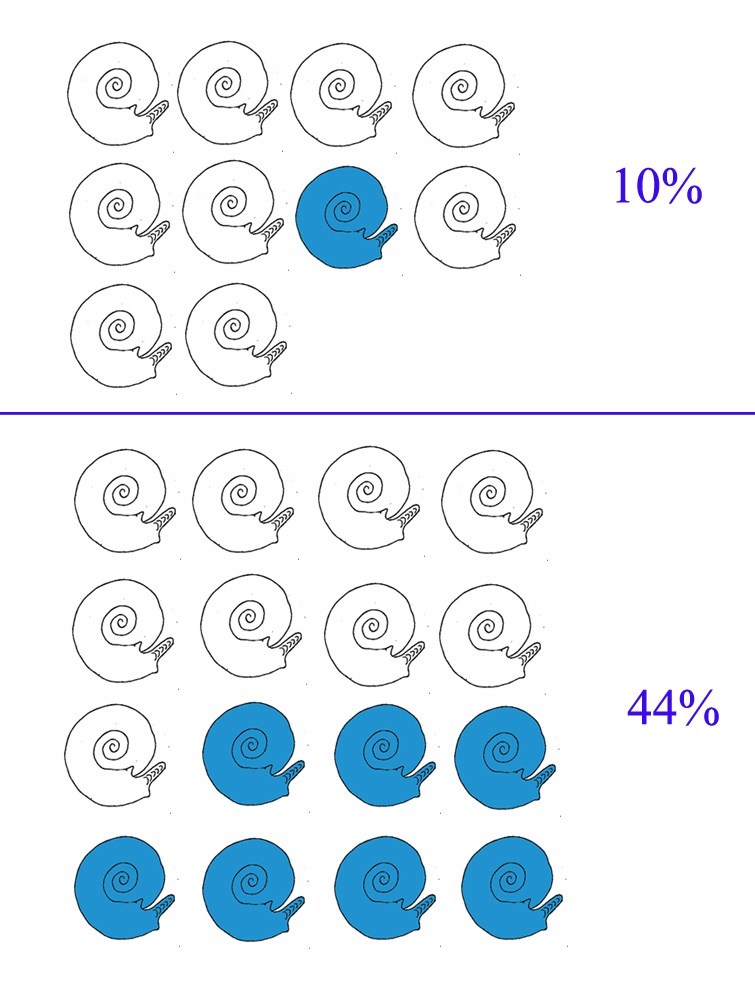
В итоге, к тому времени, когда братья и сестры того первого самца выйдут на место нереста, его «быстрорастущий генотип» будет представлен уже не одной особью, а сразу семью. То есть соотношение будет не 1 из 10 (10%), а 7 из 16 (44%). И ничего страшного, что самого этого мутанта уже и не будет в живых, эволюции до этого нет дела, главное, что сохранился его генотип. Понятно, что 7 самцов успеют спариться с большим числом самок. Их дети также быстро вырастут и через несколько поколений быстрорастущие аммониты физически вытеснят медленнорастущих. В реальных условиях этот процесс может занять всего лишь несколько десятков лет.
Но почему самки не ускоряют свое развитие так же, как самцы, ведь это выгодно и им тоже? На самом деле ускоряются и они, но до определенного предела. Они бы и рады начать размножаться как можно раньше, но формирование яиц – процесс более сложный, чем формирование мужских половых клеток. Кроме того, им необходимо построить более крупную раковину для хранения этой икры как минимум до нереста, а может быть, если у аммонитов было яйцеживорождение, то и для вынашивания оплодотворенных яиц в течение довольно длительного времени. Конечно, у самок был путь минимизации числа яиц, но «ленивые» самки, формировавшие мало яиц, скорее всего проигрывали, так как у их потомков практически не было шансов дожить до взрослого состояния – их было слишком мало, а хищников вокруг - много.
Подобную картину мы видим у современных кальмаров. Самцы у многих видов достигают зрелости уже через год после рождения и спариваются с самками старших поколений, а родившиеся одновременно с ними самки созревают на один-два сезона дольше. На это накладывается еще и время рождения, сезонность, о которой я говорил выше. Возможно, что у аммонитов с их раковинами процесс созревания самок растягивался на более долгий срок, чем аналогичный процесс у кальмаров, которым не нужно строить столь мощную конструкцию и тратить на это свои ресурсы.
Так что с микроконхами, «миниконхами» и их происхождением все более-менее понятно. Это самцы, родившиеся в разное время года, росшие при разной температуре воды и разном количестве пищи. Также и макроконхи с "мегаконхами" всего лишь самки, выросшие в разных условиях. Их рост был ограничен моментом полового созревания, а оно, в зависимости от окружающей среды, могло наступить значительно раньше или позже.
Но могли ли существовать настоящие «мегаконхи», аммониты, рост которых продолжался в течение всей их жизни? Возможно, что такие виды и существовали, но доказательств этого пока нет. А.Н. Иванов, автор термина «мегаконхи» сам на множестве примеров показал, что те или иные признаки остановки роста у мегаконхов есть.
Но даже если таких видов с бесконечным ростом среди аммонитов не было, отдельные особи аммонитов вполне могли расти неограниченно долго. Дело в том, что в биологии существует понятие «паразитической кастрации». Некоторые паразиты разрушают или угнетают половые железы организма хозяина для того, чтобы перехватить все ресурсы, направляемые животным на размножение. Особенно заметную выгоду эта стратегия приносит в том случае, если паразит живет в организме животного, погибающего после первого и единственного в жизни размножения. В таком случае, лишенный возможности размножаться организм-хозяин будет жить значительно дольше, чем все его сверстники и паразит будет процветать в его теле. А так как рост головоногих моллюсков тесно связан с их размножением, аммонит, которые никогда не созреет, никогда не прекратит расти. Случаи гигантизма, вызванного «паразитической кастрацией», известны у современных гастропод, вполне возможно, что такое может быть и у головоногих. Так что среди аммонитов вполне могли быть и настоящие «мегаконхи» - аммониты, растущие всю жизнь и так никогда и не становившиеся взрослыми по вине паразитов. Но это были редкие исключения, большинство же «мегаконхов» - макроконхи – обычные или необычно крупные самки аммонитов.
Литература:
1. Иванов А. Н. Поздний онтогенез аммонитов и его особенности у микро-‚ макро- и мегаконхов // Сб. науч. тр. Яросл. пед. ин-та. 1975. Вып. 142. С. 5-28. 4.
2. Matyja B.A. (1986) - Developmental polymorphism in Oxfordian ammonites; Acta Geol. Polon., 36 (1-3): 37-68, 4 pis.
3. Matyja B.A. (1994) "Developmental polymorphism in the Oxfordian ammonite subfamily Peltoceratinae"
4. Захаров Ю. Д. Проблема полового диморфизма ископаемых цефалопод...- Проблемы филогении и систематики. Владивосток, 1969, стр. 108—129 (Дальневост. геол. институт).
5. Callomon, J. H. (1955): The ammonite succession in the Lower Oxford Clay and Kellaway's Beds at Kidlington, Oxfordshire, and the Zones of the Callovian Stage. - Phil. Trans. royal Soc. London, B, 239
6. Несис К.Н. (2005) Головоногие: умные и стремительные. — М.: Издательство «Октопус», 2005. — 208 с
Ключевые слова (тэги): аммониты, макроконхи, микроконхи, мегаконхи, диморфизм, Ammonites, миниконхи, Microconchs, Macroconchs, Megaconchs
 Добавить в избранное
Добавить в избранное
|
Постоянный адрес публикации: |
HTML-ссылка на публикацию: |
Публикация создана 2 августа 2012 года
Если вы хотите оставить комментарий,
вам нужно зарегистрироваться (или авторизоваться, если вы уже зарегистрированы)
Комментарии:
|
<a href="http://ammonit.ru/text/615.htm#92332">http://ammonit.ru/text/615.htm#92332</a> |
|
|||||
Спасибо за познавательную публикацию! | ||||||
|
комментарий 1 уровня |
Комментарий создан 2 августа 2012 года в 01:08:22 |
|||||
 Добавить в избранное
Добавить в избранное 


|
<a href="http://ammonit.ru/text/615.htm#92334">http://ammonit.ru/text/615.htm#92334</a> |
|
|||||
Интересная публикация!
| ||||||
|
комментарий 1 уровня |
Комментарий создан 2 августа 2012 года в 01:36:07 |
|||||

|
<a href="http://ammonit.ru/text/615.htm#92378">http://ammonit.ru/text/615.htm#92378</a> |
|
|||||
Согласен, но в разных видах и популяциях аммонитов число "миниконхов" и микроконхов разное, есть и такие популяции, в которых их примерно поровну. Есть вообще мелкие популяции, с доминированием "миниконхов", Матый такие упоминал. У нас в Москве Kachpurites значительно мельче, чем на Черемухе, к примеру.
| ||||||
|
комментарий 2 уровня |
Комментарий создан 2 августа 2012 года в 09:41:57 |
|||||

|
<a href="http://ammonit.ru/text/615.htm#92421">http://ammonit.ru/text/615.htm#92421</a> |
|
|||||
Размеры кашпуритесов у нас и на Черемухе скорее всего - географические особенности популяций.
| ||||||
|
комментарий 3 уровня |
Комментарий создан 2 августа 2012 года в 13:25:03 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92361">http://ammonit.ru/text/615.htm#92361</a> |
|
|||||
С удовольствием прочитал обе части, замечательно и грамотно все рассказано.
| ||||||
|
комментарий 1 уровня |
Комментарий создан 2 августа 2012 года в 07:37:53 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92382">http://ammonit.ru/text/615.htm#92382</a> |
|
|||||
Вопрос формирования диморфных пар на самом деле очень сложный. Дискуссия о Craspedites в комментариях к первой части тому пример.
| ||||||
|
комментарий 2 уровня |
Комментарий создан 2 августа 2012 года в 10:29:03 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92386">http://ammonit.ru/text/615.htm#92386</a> |
|
|||||
Спасибо, за такой подробный ответ! Теперь окончательно стало все понятно, интересные вещи! | ||||||
|
комментарий 3 уровня |
Комментарий создан 2 августа 2012 года в 10:34:33 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92451">http://ammonit.ru/text/615.htm#92451</a> |
|
|||||
"Иногда аммониты различаются еще сильнее, как например Микроконх и макроконх (Aspidoceras cf. karpinskii и Sutneria eumela) . Но тут их объединили потому что они явно из одного семейства и живут вместе, при этом один явный микроконх, другой явный макроконх"
| ||||||
|
комментарий 3 уровня |
Комментарий создан 2 августа 2012 года в 18:12:11 |
|||||

|
<a href="http://ammonit.ru/text/615.htm#92393">http://ammonit.ru/text/615.htm#92393</a> |
|
|||||
Спасибо!
| ||||||
|
комментарий 1 уровня |
Комментарий создан 2 августа 2012 года в 10:50:05 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92394">http://ammonit.ru/text/615.htm#92394</a> |
|
|||||
Там по тексту "мужские яйцеклетки" затесались... | ||||||
|
комментарий 1 уровня |
Комментарий создан 2 августа 2012 года в 10:50:50 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92417">http://ammonit.ru/text/615.htm#92417</a> |
|
|||||
Неточность формулировок, исправлю )) | ||||||
|
комментарий 2 уровня |
Комментарий создан 2 августа 2012 года в 13:00:51 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92404">http://ammonit.ru/text/615.htm#92404</a> |
|
|||||
Трудно сказать,что интересней-публикация или комментарии... Хорошая тема! | ||||||
|
комментарий 1 уровня |
Комментарий создан 2 августа 2012 года в 12:01:26 |
|||||

|
<a href="http://ammonit.ru/text/615.htm#92449">http://ammonit.ru/text/615.htm#92449</a> |
|
|||||
С микро и макроконхами действительно все более, менее ясно,спасибо за объяснения.К тому же быстрое созревание объясняет огромное количество головоногих в те времена.Идея полового диморфизма звучит более доказательно.А вот по мега и миниконхам очень понравилось предположение Mystriver.Ведь действительно леса Крыма съедает раз в 5-6 лет непарный шелкопряд почти под чистую...Это не головоногий и даже не морской обиталь,а всего лишь бабочка вредитель)))Но!!!В годы, когда его потомство не массово,особенно после достаточно сильных холодов, самки этих бабочек вырастают огромного размера(почти в 2 раза больше чем в обычные годы),причем они даже летают с трудом.Откладывают огромное количество яиц и гусеницы съедают весь лес под чистую!Самцы же остаются прежних размеров.Так что я думаю в годовых колебаниях температуры или кормовой базы в морях того времени и кроется ответ на всплески развития среди аммонитов мини и мегаконхов.Поэтому их и не так много, как микро и макроконхов. | ||||||
|
комментарий 1 уровня |
Комментарий создан 2 августа 2012 года в 17:19:25 |
|||||

|
<a href="http://ammonit.ru/text/615.htm#92458">http://ammonit.ru/text/615.htm#92458</a> |
|
|||||
Отличная статья! теперь понятно что мой Peltoceras - быстро выросший микроконх | ||||||
|
комментарий 1 уровня |
Комментарий создан 2 августа 2012 года в 19:05:15 |
|||||

|
<a href="http://ammonit.ru/text/615.htm#92462">http://ammonit.ru/text/615.htm#92462</a> |
|
|||||
Скорее наоборот долго росший )) Аммонит если взрослел быстро, то был небольшим, а если он крупный, то скорее всего наоборот рос немного дольше. Хотя они могли с мелкими микроконхами и одинаковое время расти но в разных условиях, у кальмаров и такое бывает.
| ||||||
|
комментарий 2 уровня |
Комментарий создан 2 августа 2012 года в 19:11:34 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92525">http://ammonit.ru/text/615.htm#92525</a> |
|
|||||
Интересно и много нового!
| ||||||
|
комментарий 1 уровня |
Комментарий создан 2 августа 2012 года в 22:07:48 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92623">http://ammonit.ru/text/615.htm#92623</a> |
|
|||||
сезоны были, климатические пояса были
| ||||||
|
комментарий 2 уровня |
Комментарий создан 3 августа 2012 года в 15:49:50 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92640">http://ammonit.ru/text/615.htm#92640</a> |
|
|||||
Спасибо, значит зима и лето, хоть и мягкие, но все же были. | ||||||
|
комментарий 3 уровня |
Комментарий создан 3 августа 2012 года в 20:41:48 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92646">http://ammonit.ru/text/615.htm#92646</a> |
|
|||||
у нас были примерно субтропики, а в Арктике и сезонные льды бывали | ||||||
|
комментарий 4 уровня |
Комментарий создан 3 августа 2012 года в 20:59:57 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92647">http://ammonit.ru/text/615.htm#92647</a> |
|
|||||
Получается. что аммониты обитали по климатическим зонам, в зависимости от широты, и еще в зависмомти от глубины океанов? | ||||||
|
комментарий 5 уровня |
Комментарий создан 3 августа 2012 года в 21:05:54 |
|||||
|
<a href="http://ammonit.ru/text/615.htm#92681">http://ammonit.ru/text/615.htm#92681</a> |
|
|||||
да | ||||||
|
комментарий 6 уровня |
Комментарий создан 3 августа 2012 года в 23:10:25 |
|||||